

- 3.28 MB
- 2022-04-29 14:47:53 发布

- 1、本文档共5页,可阅读全部内容。
- 2、本文档内容版权归属内容提供方,所产生的收益全部归内容提供方所有。如果您对本文有版权争议,可选择认领,认领后既往收益都归您。
- 3、本文档由用户上传,本站不保证质量和数量令人满意,可能有诸多瑕疵,付费之前,请仔细先通过免费阅读内容等途径辨别内容交易风险。如存在严重挂羊头卖狗肉之情形,可联系本站下载客服投诉处理。
- 文档侵权举报电话:19940600175。
'基因修饰在人源化肝脏小鼠建模中的应用
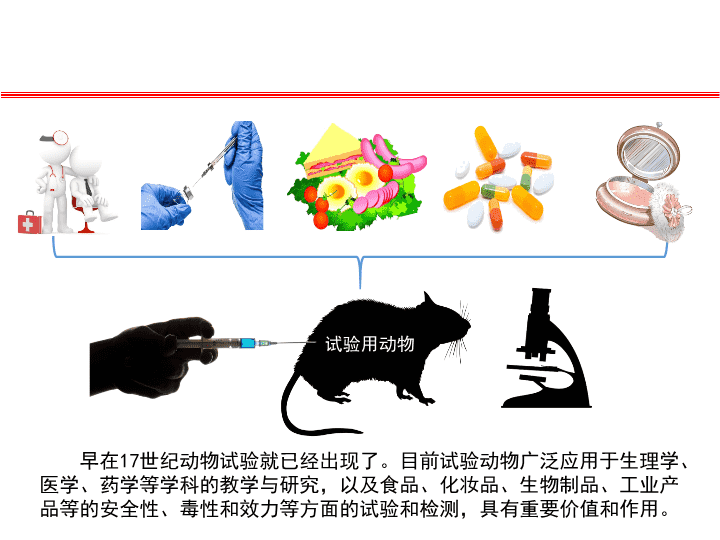
早在17世纪动物试验就已经出现了。目前试验动物广泛应用于生理学、医学、药学等学科的教学与研究,以及食品、化妆品、生物制品、工业产品等的安全性、毒性和效力等方面的试验和检测,具有重要价值和作用。试验用动物
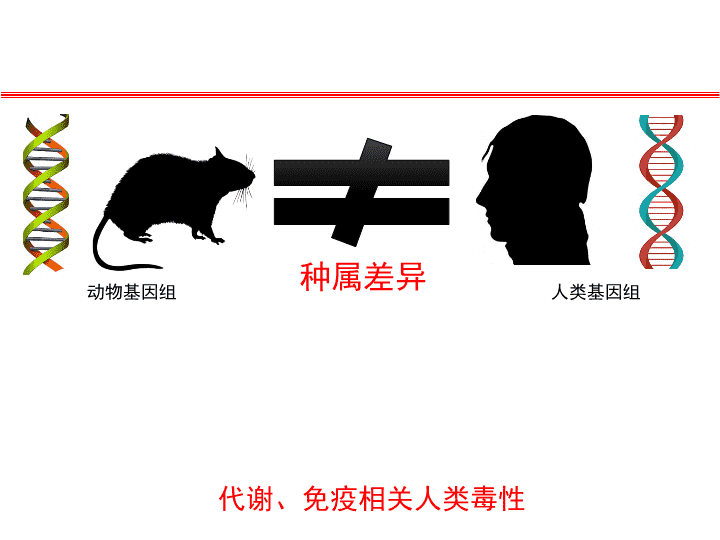
种属差异动物基因组人类基因组代谢、免疫相关人类毒性
试验动物作为预测工具如不能完全准确预测人体可能会出现的情况,就如同战地扫雷,一旦出现误判,代价很有可能将是巨大的!危险安全
那么能不能构建一种临床前更贴近人肝代谢、预测性更一致性的试验动物模型,将可能发生的人类肝毒性提前筛查出来,降低无谓成本,减少临床损失呢?这将具有极其重大的意义和价值!人源化肝脏小鼠模型由此成为研究热点
现代生物医药领域研究模式基因修饰技术ES打靶基因修饰技术TALEN基因修饰技术CRISPR/Cas9基因修饰技术TetraOneTM基因修饰技术基因修饰动物建模
免疫缺陷小鼠模型的构建类别序号名称基因信息特异性免疫非特异性免疫免疫缺陷小鼠1NudeFoxn1nu无胸腺,无T,B↓正常2SCIDPrkdcscid无T,无B正常3NOD-SCIDNOD/Prkdcscid无T,无BNK↓补体↓4Rag2Rag2-/-T、B功能缺失正常5IL2RGIl2rg-/-T、B部分功能缺失NK↓6F344RGRag2-/-/Il2rg-/-T、B功能缺失NK功能缺失7FPGPrkdc-/-/Il2rg-/-T、B功能缺失NK功能缺失8NOGNOD/Prkdcscid/Il2rgtm1Sug无T,无BNK、补体功能缺失
人源化肝脏小鼠模型的构建工程基因片段uPA基因片段Fah基因片段HSVtk/GCV自杀系统Tet-on/uPA自杀系统其他移植用人类细胞肝细胞造血干细胞胸腺细胞其他
(1)uPA-Rag2小鼠(Dandri,2001年,德国)uPA全称尿激酶纤维蛋白溶酶原活化子(Urokinase-typePlasminogenActivator),当白蛋白启动的uPA基因片段在小鼠肝细胞内表达时,纤维蛋白溶酶原会激活纤维蛋白酶,诱发粗面内质网发生裂解损伤,最后肝细胞损伤并逐渐消亡。Rag2全称重组激活基因2(Recombination-ActivatingGene2),在VDJ基因重排和淋巴细胞发育中起关键性作用。Rag2-/-小鼠为C57BL/6J品系背景,T、B淋巴细胞早期发育严重停滞,外周血没有成熟的循环T、B淋巴细胞,非特异性免疫收影响小。Rag2小鼠uPA基因肝细胞人源化率仅有15%,适合病毒学研究,但无法满足代谢研究需求!!
(2)uPA-SCID小鼠(Tateno,2004年,日本)SCID小鼠全名联合缺陷免疫小鼠(severecombinedimmunedeficiency),其第16对染色体隐性基因突变,纯合子的淋巴细胞抗原受体基因VDJ区域的重组酶活性异常,基因重排无法正常进行,影响B、T细胞的正常分化,外周血白细胞总数减少,但粒细胞、巨噬细胞、NK细胞、LAK细胞正常。uPA-SCID小鼠相比uPA-Rag2小鼠具有更高的人源化比率,其肝细胞替换可达50%,运用免疫抑制剂后可提升至70%,基本满足代谢和毒性研究需求,许多学者利用该模型进行了多种药物研究。SCID小鼠uPA基因
持续性自发肝损;出血性疾病导致死亡;高血补体浓度(C3)肾病;生殖障碍;移植窗较小(出生后2W内移植);存在非特异性免疫,需药物抑制免疫。名称基因信息特异性免疫非特异性免疫肝损伤uPA-SCIDuPA/Prkdcscid无T无B正常uPA(2)uPA-SCID小鼠(Tateno,2004年,日本)
(3)Fah小鼠(Grompe,1993年,波兰)Fah全称延胡索酰乙酰乙酸水解酶基因(fumarylacetoacetatehydrolase),Fah-/-小鼠酪氨酸代谢受阻,出现Ⅰ型酪氨酸血症,毒性产物损伤肝、肾。2-硝基-4-三氟甲苯-1,3环乙烷碘(NTBC)可以抑制酪氨酸降解通路中4-对羟基苯丙酮酸双氧化酶(4-HPPD)活性,减少毒性代谢产物积聚,起到治疗作用。NTBC治疗维持具有免疫系统,不适合人源化组织和细胞移植
Fah/nudeFah/Rag1Fah/Nod/Scidll2rg基因是白细胞介素2受体的gamma链(Il-2Rγc,又称CD132),是具有免疫功能的细胞因子Il-2、Il-4、Il-7、Il-9、Il-15和I-21的共同受体亚基。L2rg-/-小鼠机体免疫功能严重降低,尤其是NK细胞的活性几乎丧失。名称基因信息特异性免疫非特异性免疫肝损伤FRGFah-/-/Rag2-/-/Il2rg-/-T、B功能缺失NK↓Fah-/-(4)FRG小鼠(Azuma,2007年,波兰)
持续性的自发肝脏;自发性Ⅰ型酪氨酸血症突出,易诱发肝癌;需要药物NTBC控制肝损伤程度,易对试验结果造成影响;小鼠存活率不高,饲养时易死亡。(4)FRG小鼠(Azuma,2007年,波兰)FRG小鼠在性状和效果上都优于Fah-/-小鼠,其肝细胞替换率达80%以上,后经其他学者改进探索,可达95%左右。该模型应用于代谢和病毒等多个领域研究,有重要意义。
名称基因信息特异性免疫非特异性免疫NOGNOD/Prkdcscid/Il2rgtm1Sug无T,无BNK、补体功能缺失(5)NOG、NSG小鼠(Ito,2002年,日本)NOG小鼠由日本实验动物研究所的Ito培育而成。通过杂交NOD/Scid和Il2rg-/-小鼠而来,其T、B细胞皆缺失,NK细胞功能缺陷。与NOD/Scid小鼠相比,NOG小鼠的人体细胞和组织移植存活率显著提高,同时能够植入更高比例的正常或癌变人类细胞和组织。NOG、NSG是类似的并行品系,源于日本为NOG(NOD/Shi-scid/Il-2Rγcnull),源于美国为NSG(NOD/scid/γC−/−/SzJ)。NOG小鼠在免疫缺陷模型、肿瘤模型、异种移植模型、传染病模型领域有大量应用。
名称基因改变特异性免疫非特异性免疫肝损伤TK-NOGTK/NOD/Prkdcscid/Il2rgtm1Sug无T,无BNK、补体功能缺失HSVtk/GCV系统(6)TK-NOG小鼠(Hasegawa,2011年,日本)单纯疱疹病毒胸苷激酶/更昔洛韦(HSVtk/GCV)系统是分子生物学基因修饰技术中运用十分广泛的自杀系统,GCV本身无毒,但是在转染了HSVtk基因的细胞中可被磷酸化,成为单磷酸盐,再经内源性细胞激酶作用可转化为二磷酸和三磷酸盐,对细胞有高毒性。♂♀♀NOGTK-NOG
具有人肝等效的酶表达及生物功能;具有较高立体组织结构;人源化程度高(≥90%);生存率高,移植时段宽裕,可重复利用;无需外源性药物维持性状;可生殖传代(雌性可与NOG小鼠生育);在代谢、毒性、病毒、微生物方面大量应用2011-2017,美日有诸多研究和数据。无商业化成品供应,国内来源受限!(6)TK-NOG小鼠(Hasegawa,2011年,日本)
(7)uPA-NOG小鼠(Gutti,2014年,美国)名称基因改变特异性免疫非特异性免疫肝损伤uPA-NOGuPA/NOD/Shi-scid/Il2rgnull无T,无BNK、补体功能缺失uPAuPA-NOG小鼠是在NOG小鼠基础上通过基因修饰加入uPA基因培育而来,目前应用相对较少,有报导的文献显示仅应用于病毒学研究,对肝脏人源化指数没有很高要求,仅10%左右。(8)FRGN小鼠(Wilson,2014年,美国)名称基因改变特异性免疫非特异性免疫肝损伤FRGNFah−/−/NOD/Rag2−/−/Il2rg−/−无T,无BNK、补体功能缺失Fah−/−FRGN小鼠是在NOD小鼠基础上敲除Fah、Rag2、Il2rg基因培育而来,目前应用相对较少,文献报导肝脏人源化率可达80%,并且在免疫重建上也有一定应用趋势。
请多多指正,谢谢!
比较解剖学
皮肤及其衍生物表皮-复层上皮组织(胚胎外胚层)皮肤:真皮-结缔组织(中胚层)表皮衍生物:表皮外骨骼(角质鳞、羽、毛。喙、爪、蹄、指甲、空角)和腺体真皮衍生物:骨质鳞(硬鳞、圆鳞、栉鳞)鳍条、实角、骨板等
各类脊椎动物皮肤及其衍生物的比较文昌鱼:表皮-单层柱状上皮;真皮-胶状结缔组织圆口类:表皮-多层上皮;真皮-胶状结缔组织鱼类:同圆口类体背被鳞-楯鳞、硬鳞、圆鳞和栉鳞
楯鳞:板鳃鱼特有,由基板(埋在真皮内,有神经和血管)和棘突(向后伸于皮肤外,外被釉质-来源于外胚层,表皮细胞所分泌)组成。硬鳞:硬鳞鱼特有(雀鳝、鲟鱼),由真皮形成骨质板;表面覆有坚硬的闪光质,鳞片多菱型,成对角线排列。圆鳞:由真皮演化来的骨质鳞,鳞前端斜插入真皮内,后端游离,呈覆瓦状排列于表皮下。栉鳞:同圆鳞,但游离端具数列锯齿状突起,见于较高等的硬骨鱼。硬鳞、圆鳞和栉鳞-来源于真皮,属骨组织和纤维质
两栖类-仅表层的1~2层细胞开始角质化,蜕皮开始从其类开始,真皮厚而致密。显示陆生脊椎动物的特征。
爬行类-角质化加深,表皮明显分为角质层和生长层,蜕皮现象特明显,指趾端出现爪。真皮薄。
鸟类-表皮薄软松,无羽的腿等部被以角质鳞。真皮薄无衍生物。
哺乳类-皮肤厚坚韧,真皮非常发达。毛为该类特有。皮肤腺异常发达。
骨骼系统硬骨形成的方式:从结缔组织经软骨再变为硬骨-软骨原骨(脊柱、肋骨、四肢骨)在结缔组织基础上,不经过软骨阶段而直接形成硬骨-膜原骨或膜性硬骨(鼻骨、额骨等)
椎体类型双凹型:椎体活动有限-鱼类、有尾两栖类和少数爬行类前凹型:椎体前端凹入而后端凸出,两椎体活动较为灵活。两栖类(多数无尾类)和多数爬行类和鸟类的第一颈椎后凹型:椎体前端凸出而后端凹入。椎体相接形成活动关节。两栖类(多数蝾螈类及无尾类一部分)少数爬行类
马鞍型:椎间活动性极大,脊索不存在。鸟类颈椎双平型:椎体前后两端扁平,椎体间垫以纤维软骨的椎间盘以减少活动摩擦。在椎间盘内仍留有少量残余脊索(髓核)哺乳类
各类脊椎动物脊柱的比较圆口类:脊索保留终身鱼类:脊柱分为躯椎和尾椎两栖类:脊柱除一般的增加坚固外,脊柱分为颈、躯、荐、尾4区。爬行类:脊柱分为颈、胸、腰、荐、尾5区。颈椎数较两栖类多,且颈椎又分化为寰椎和枢椎,胸椎极其明显,与肋骨、胸骨相接构成胸廓
鸟类:分区与爬行类相同,但颈椎数目加多,椎体呈马鞍型,最后一个胸椎、全部腰椎和荐椎、连同前面的几个尾椎完全愈合为一个整体-综荐骨,再与腰带相接。哺乳类:分为5区,颈椎恒为7块,胸椎数目变化较大。胸骨从两栖类开始出现。胸廓是由胸椎、肋骨及胸骨借助关节、韧带连接而成。
头骨的组成脊椎动物的头骨在发生上是由下列3个来源不同的部分组合而成,及软颅、咽颅和膜颅。软颅:原始的软骨脑颅,保护脑及感官咽颅:围绕消化管的前端,保护和支持口腔、舌及鳃。在水生脊椎动物中,咽颅发达,但和脑颅关系不太密切,仅以韧带和脑颅相接。
膜颅:为一系列真皮骨骼,直接由间充质骨化形成,不经软骨阶段。覆盖在软颅上(主要是在顶部)或咽颅上,成为头骨的膜原骨部分。
消化系统消化道+消化腺--由胚胎期的原肠及其突出分行形成。管壁构造:粘膜+粘膜下层+肌肉层+浆膜牙齿:脊椎动物的牙齿与软骨鱼的楯鳞同源,全部由外和中胚层共同形成。文昌鱼和无颌类无齿,牙齿是伴着颌的出现而产生。其最初的功能:捕捉及咬着食物,进化到哺乳类:切割+刺穿+撕裂+研磨。现代鸟类次生性缺齿。
牙齿的进化历程由同型齿-异齿型多出齿-再生齿端生齿或侧生齿-槽生齿数量多而不恒定到量少而恒定由着生部位广泛(上下颌、梨骨、腭骨、副碟骨)到仅生于上下颌。
舌和口腺文昌鱼无舌。无颌类有舌,适应半寄生,舌象活塞,以舌端角质齿挫破皮肤吸食其血肉。有特殊的口腺(抗凝血),鱼类有舌,但无舌内肌肉(前后挪动帮助吞食)。空腔内无口腺无尾两栖类以上,舌有舌内肌,能自由伸缩。两栖类开始有口腺爬行类有鳞类的舌活动性更大。口腺发达(唇腺、腭腺、舌腺和舌下腺),分泌物湿润食物、粘捕食物。毒腺是变态的口腺。鸟类的舌硬,表面被角化的上皮。食谷的口腺发达,但不含消化酶的唾液,仅湿润食物(燕雀类唾液中有消化酶)哺乳类舌富肌肉,其背粘膜具乳头(内有味蕾)。口腺发达(耳下腺、颌下腺和舌下腺,兔另有眶下腺),唾液中有消化酶。
咽鱼类,水与食物共同入口,到咽部水由鳃裂流出,同时在鳃部行气体交换,食物入食道。自两栖类内鼻孔出现,空气由鼻孔---空腔到肺,食物由口入食道,彼此在口腔交叉后前行。哺乳类的内鼻孔后移,硬腭之后延伸出软腭,空气通道和食物通道在咽部形成交叉。
食道和胃文昌鱼和无颌类无食道和胃分化。鱼类出现胃。爬行类以上,随着颈部的延长,相应有很长的食道。鸟类的食道在中部扩大成嗉囊。胃的形态、位置随体型、身体位置和食性的不同有很大变化。鸟类胃有腺胃和肌胃的分化,反刍哺乳类胃分瘤胃、网胃、瓣胃和皱胃。
肠在进化中其一方面增加分化程度,一方面增加消化吸收面积。文昌鱼、无颌类、鱼类的肠尚无明显分化,两栖类有大小肠分化,但大肠很短。爬行类在大肠和小肠间第一次出现了盲肠。鸟类有一对盲肠,在草食性和某些杂食性的鸟类身上,盲肠非常发达。哺乳类的小肠分化为十二指肠、空肠和回肠。大肠分化为结肠和直肠。草食性兽类的盲肠很大。
肠增加肠的吸收面积有多种方式:七鳃鳗沿肠管有螺旋状得粘膜褶伸入肠腔内,称盲沟。依靠螺旋瓣来增加肠的面积是较古老的一种方式,如鲨鱼、银鲛、非洲肺鱼和鲟鱼等。现代陆生四足类都不再保留螺旋瓣结构。硬骨鱼类没有螺旋瓣,但在某些种类如鲈鱼具有幽门盲囊。多数脊椎动物是靠增加肠的长度来增加面积。植食性的肠较长,肉食性的较短。增加小肠的吸收面积还靠小肠粘膜向管内腔突起所形成的皱褶和绒毛。
泄殖腔:是肠的末端略为膨大处,输尿管和生殖管窦开口于此腔,因之此腔是粪、尿和生殖细胞共同排出的地方,以单一的泄殖腔孔开口于体外。软骨鱼、两栖类、爬行类、鸟类和哺乳类中的单孔类皆具泄殖腔。泄殖腔与泄殖窦
泄殖窦:是输尿管和生殖管汇合的略为膨大处,以泄殖孔开口于体外,肠管单独以肛门开口于外。圆口类、硬骨鱼、哺乳类皆属此类。其肛门和泄殖孔皆是分别开口的,雌性灵长类的排泄和生殖管道也分别开口的。泄殖腔与泄殖窦
颌弓和脑颅相接的方式两接型:上颌骨以韧带与头骨相连,舌弓又通过舌颌骨以韧带连头骨与上下颌。-原始软骨鱼。舌接型:以舌颌骨作悬器,将上下颌连与头骨。多数软骨鱼和硬骨鱼自接型:舌颌骨移入中耳,不再作为颌额悬器,上颌腭方软骨直接与脑颅愈合,其上的方骨与下颌的关节骨成关节。陆生脊椎动物大多属此。颅接型:上颌腭方软骨直接与脑颅愈合,方骨和关节骨变为中耳听小骨,下颌的齿骨直接连脑颅。-哺乳类
动脉系统脊椎动物主要的动脉在各纲大致相同,皆包括:1、腹大动脉由心脏发出,沿消化管的腹面向前走;2、背大动脉为一对位于消化管背面的纵行血管,向后走行一段距离后,在愈合成单条的背大动脉,发出分支到全身各部;3、6对动脉弓连接腹大动脉和背大动脉
各纲动物动脉系统的演变主要是动脉弓的变化,这种改变是由鳃呼吸进化为肺呼吸而引起的。鱼类:每一动脉弓分为两部分,一部是入鳃动脉,一部是出鳃动脉,两者之间有毛细血管。前者由腹大动脉运送血液入鳃,在鳃部毛细血管进行气体交换,再由出鳃动脉把富氧血集中到大动脉根。
无尾两栖类:蝌蚪期入硬骨鱼,保留4对入鳃和出鳃动脉,代表第三至六对动脉弓。变态后,营肺呼吸,鳃消失,全部动脉弓不再以毛细血管相连,其中第三对动脉弓形成颈动脉,第四对形成体动脉,左右体动脉向下弯曲成弓后合成背大动脉,第六对动脉弓转变为肺皮动脉,分支到肺和皮肤中。原胚胎期的第一、二、五对动脉弓消失。有尾两栖类仍保留退化的第五对动脉弓,肺动脉和体动脉弓间由动脉导管相通。
爬行类:具第三、四、六对动脉弓,一般爬行类的成体动脉导管和颈动脉管皆关闭,但低等的蜥蜴类的第三对和第四对动脉弓之间的颈动脉管还保留;某些蛇和龟鳖类在第四和第六对动脉弓之间还有动脉导管连接。爬行类的肺动脉、左、右体动脉3个主干分别由心脏发出,其中肺动脉和左体动脉弓由心室的右侧发出,右体动脉弓由心室的左侧发出。
鸟类:成体左侧的体动脉弓完全退化,只保留右体动脉弓;哺乳类:仅保留左体动脉弓。
排泄系统不同的排泄物是通过不同的途径排出体外。除一部分通过皮肤随汗排出,一部分二氧化碳和水通过肺排出外,绝大多数代谢废物(尿素、尿酸、铵盐和水分)是随血液通过肾脏以尿的形式排出。脊椎动物的排泄系统包括肾脏、输尿管、膀胱和尿道。
肾脏的类型肾脏由中胚层的中节形成的生肾节组成。无羊膜肾脏的发生要经过前肾(胚胎期)和背肾(成体)两个阶段;羊膜类动物需经过3个阶段:即前肾、中肾和后肾阶段。脊椎动物的肾脏总共可以分为全肾、前肾、中肾、背肾和后肾5种类型。
输尿管和膀胱输尿管七鳃鳗的中肾管即输尿管,只有输尿作用,与生殖无关。软骨鱼另外形成多条副肾管输尿,而中肾管作为输精管。硬骨鱼情况特殊,中肾管仅作输尿之用。两栖类的中肾管即输尿管,在雄性兼做输精之用。羊膜动物发展出后肾,其输尿管即为后肾管。部分爬行类和鸟类因缺少膀胱,输尿管直接通入泄殖腔,而在哺乳类则输尿管先开口与膀胱,再以尿道通体外。
膀胱圆口类、软骨鱼、部分爬行类和鸟类全无膀胱,其它脊椎动物皆有膀胱。膀胱分3种类型:导管膀胱:由中肾管后端膨大形成,见于硬鳞鱼,硬骨鱼,但少数硬骨鱼(比目鱼科)无膀胱。泄殖腔膀胱:由泄殖腔腹壁突出而成,故中肾与膀胱不直接发生联系。如肺鱼、两栖类和哺乳类的单孔类。尿囊膀胱:有胚胎时期尿囊柄的基部膨大而成。如少数爬行动物(龟鳖、、部分蜥蜴类)
'
您可能关注的文档
- 最新基因身份证分析课件PPT.ppt
- 最新基因诊断技术的应用课件PPT.ppt
- 最新基因表达调控ppt课件课件PPT.ppt
- 最新基因组学技巧在致病基因发觉课件PPT.ppt
- 最新基因工程菌的发酵课件PPT.ppt
- 最新基因工程第2章 酶学基础(2dna连接酶)课件PPT.ppt
- 最新基因工程及其应用课件PPT课件.ppt
- 最新基因对性状的控制课件PPT课件.ppt
- 最新基因功能研究技术课件PPT.ppt
- 最新基于阈值和区域生长法的图像分割方法课件PPT.ppt
- 最新基于素质模型的结构化面试三只眼(精简版)[PPT课件]课件PPT.ppt
- 最新基于热电偶温度传感器的动态温度实时测量记录系统设计.课件PPT.ppt
- 最新基于机器视觉技术发动机曲轴轴瓦分级检测系统应用研究课件PPT.ppt
- 最新基于听感知的 音频信息隐藏研究课件PPT.ppt
- 最新基于决策树改进CART算法的ANFIS结构辨识技术-(1)课件PPT.ppt
- 最新基于FPGA的DSP开发设计精要课件PPT.ppt
- 最新基于DSP的汽车运行状况监测系统硬件设计课件PPT.ppt
- 最新培达血管外科课件PPT.ppt